포유류의 진화
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
포유류의 진화는 약 3억 2천만 년 전 단궁류의 출현으로 시작되어, 젖샘과 중이의 특징을 가진 포유류로 발전했다. 초기 포유류는 야행성 생활을 하며 털, 높은 신진대사, 큰 뇌를 발달시켰고, 페름기-트라이아스기 대멸종 이후 아르코사우루스류와의 경쟁에서 살아남아 다양한 생태적 지위를 차지했다. 현생 포유류는 단공류, 유대류, 유태반류로 분류되며, 각 그룹은 독특한 번식 방식과 특징을 보인다. 화석 연구와 분자 계통학을 통해 포유류의 진화 과정을 추적하고 있으며, 털, 젖 분비, 뇌 발달 등 다양한 측면에서 진화적 변화를 관찰할 수 있다.
더 읽어볼만한 페이지
- 포유류학 - 고래학
고래학은 고래류에 대한 연구를 수행하는 학문 분야이며, 고대부터 관찰 기록이 존재하고, 환경 문제와 수족관 돌고래 쇼의 인기로 연구가 활발하게 진행되고 있다. - 포유류학 - 세계의 포유류 종
세계의 포유류 종은 1982년 초판 발행 이후 지속적으로 개정 및 증보되어 현재 5,416종 이상을 포함하고 있으며, 돈 E. 윌슨과 디앤 M. 리더가 오랫동안 편집을 담당했고, 가장 최근 판은 온라인으로 운영되며 제4판 발행이 예정된 포유류 종 목록 책이다. - 척추동물의 진화 - 다윈의 핀치
다윈의 핀치는 갈라파고스 제도에 서식하며 자연 선택에 의한 진화의 대표적인 사례로 꼽히는 핀치새들로, 다양한 환경에 적응하여 부리 모양과 크기가 다른 여러 종으로 분화되었고, 뼈 형태 형성 단백질 4(BMP4) 및 칼모듈린 유전자와 같은 분자적 기반 연구를 통해 부리 형태 변이의 유전적 메커니즘이 밝혀지고 있다. - 척추동물의 진화 - 조류의 진화
조류의 진화는 수각류 공룡에서 시작되어 깃털 발달과 꼬리 퇴화를 거쳐 현대 조류의 특징을 갖추게 되었으며, 현재는 다양한 환경 변화에 직면해 있다. - 고생물학 - 대량절멸
대량절멸은 지구 역사에서 생물 다양성이 급격히 감소하는 현상으로, 5번의 주요 멸종과 현재 진행 중인 홀로세 멸종을 포함하여 지구 생태계에 큰 영향을 미치며, 특히 홀로세 멸종은 인간 활동으로 인해 멸종 속도가 자연 멸종 속도보다 훨씬 빠르게 증가하고 있다. - 고생물학 - 규화목
나무가 땅에 묻혀 광물질에 의해 화석화된 규화목은 일본과 미국 등지에서 발견되며, 특히 일본에서는 다양한 종류가 천연기념물로 지정되어 보호받고 있다.
2. 포유류의 기원과 진화
포유류는 중생대 말기에 등장했는데, 이 시대에는 현재의 캥거루와 같은 유대류가 모두 나무 위에서 생활하고 있었다. 백악기 말이 되면서 이들이 출현하게 되었고, 이 시대에 식충류도 출현하여 모두 수상생활을 하고 있었다. 그러다가 초원이 퍼져감에 따라 평지에서 사는 포유류가 증가했다.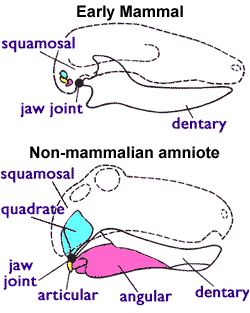
현존하는 포유류 종은 암컷의 젖샘 존재 여부로 식별할 수 있지만, 젖샘과 기타 연조직 특징은 화석에서 보이지 않기 때문에 화석을 분류할 때는 다른 특징이 필요하다.
고생물학에서 사용할 수 있는 그러한 특징 중 하나는 모든 현존하는 포유류(단공류 포함)가 공유하지만 초기 트라이아스기 수궁류에서는 나타나지 않는 특징으로, 그림 1(오른쪽)에 나와 있다. 즉, 포유류는 다른 모든 양막류가 먹는 데 사용하는 두 개의 뼈를 청각에 사용한다. 초기 양막류는 관절뼈(아래턱 뒤쪽의 작은 뼈)와 네모뼈(위턱 뒤쪽의 작은 뼈)로 구성된 턱 관절을 가지고 있었다. 양서류, 거북이, 도마뱀, 뱀, 악어류, 공룡(조류 포함), 어룡, 익룡 및 수궁류를 포함한 모든 비포유류 사지동물이 이 시스템을 사용한다. 그러나 포유류는 치골(이를 지닌 아래턱 뼈)과 인상골(또 다른 작은 두개골 뼈)로만 구성된 다른 턱 관절을 가지고 있다. 쥐라기 시대에 네모뼈와 관절뼈는 중이의 침골과 망치뼈로 진화했다.[5][6] 포유류는 또한 이중 후두 융기를 가지는데, 다른 사지동물은 단일 후두 융기를 가진다.
1981년 케네스 A. 커맥과 그의 공동 저자들은 포유류와 초기 시냅시드 사이의 경계선을 포유류형 어금니 교합 패턴이 획득되고 치골-인상골 관절이 나타나는 지점으로 긋는 것을 주장했다. 그들의 선택은 "아래턱이 중생대 포유류의 보존될 가능성이 가장 높은 골격 요소"라는 사실에 근거했다.[7] 오늘날, 대부분의 고생물학자들은 이 기준을 충족하는 동물을 포유류로 간주한다.[39]
최초의 완전 육상 척추동물은 파충류 양막류였다. 이들의 알은 내부 막을 가지고 있어서 배아가 호흡할 수 있으면서도 수분을 유지했다. 이는 양막류가 건조한 육지에 알을 낳을 수 있게 해준 반면, 양서류는 일반적으로 물속에 알을 낳아야 했다. 최초의 양막류는 석탄기 중기에 조상인 파충형류로부터 나타난 것으로 보인다.[8]
수백만 년 이내에 두 개의 중요한 양막류 계통이 분화되었다. 단궁류는 포유류의 조상이며, 파충류는 도마뱀, 뱀, 거북, 악어, 공룡, 그리고 조류의 조상이다.[3] 단궁류와 파충류의 가장 초기의 화석(각각 ''아르케오티리스''와 ''힐로노무스'' 등)은 약 에서 전으로 거슬러 올라간다. 척추동물 화석이 석탄기 후반에는 매우 희귀하기 때문에, 기원 시기를 정확히 알기는 어렵다. 따라서 각 동물 유형의 실제 최초 출현 시기는 최초의 화석 발견보다 훨씬 이전일 수 있다.[9]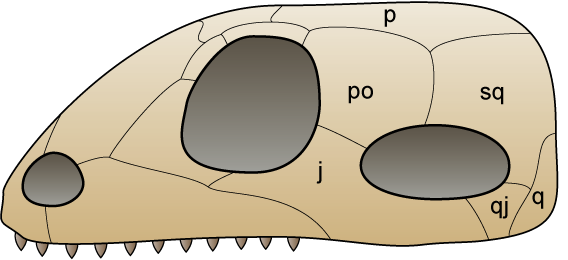
시냅스류(Synapsida) 두개골은 각 눈 뒤에 있는 구멍의 독특한 패턴으로 식별되며, 이는 다음과 같은 목적을 수행했다.
- 두개골의 강성을 희생하지 않으면서 가볍게 만들었다.
- 뼈를 덜 사용하여 에너지를 절약했다.
- 아마도 턱 근육의 부착 지점을 제공했을 것이다.
자주, 그리고 잘못되게 공룡으로 여겨지는 많은 생물체들은 파충류 계통과 사우롭시드의 일부였지만, 사실은 시냅스류였다. 여기에는 잘 알려진 ''디메트로돈''이 포함된다.[10][11]

테라프시드는 초기 단궁류인 스페나코돈티아에서 과달루페 절 중기에 기원했으며, 육상 척추동물로서 그들을 대체하여 우위를 점했다. 그들은 더 큰 측두창과 크기가 같은 앞니를 포함하여 두개골과 턱의 여러 특징에서 이전의 단궁류와 달랐다.[13]
테라프시드 계통은 여러 단계를 거쳐 페름기 후기에 키노돈의 진화를 이끌었고, 이 중 일부는 초기 포유류와 비슷해지기 시작했다:[14]
- 점진적인 뼈로 된 이차 경구개의 발달.
- 치골이 점차 아래턱의 주요 뼈가 되었다.

키노돈트(Cynodontia)는 후기 페름기에 등장한 수수류 그룹으로, 모든 포유류의 조상을 포함한다. 키노돈트의 포유류와 유사한 특징으로는 아래턱 뼈의 수 감소, 이차적인 경구개, 치관에 복잡한 패턴이 있는 볼 이빨, 그리고 두개강을 채우는 뇌가 있다.[21]
이러한 진화적 변화는 최초의 포유류(크기 약 약 10.16cm)로 이어졌다. 그들은 빠른 성장과 짧은 수명을 진화시킨 것으로 보이며, 이러한 생활사는 수많은 현대 소형 포유류에서도 발견된다.[23]
(Cynodontia:Dendrogram – Palaeos를 기반으로 함[40])
{{clade| style=font-size:90%;line-height:100%
|label1=키노돈트
|1={{clade
|1={{clade
|1=''드비니아''
|2=프로키노수키드
}}
|label2=에피키노돈트
|2={{clade
|1=''트리낙소돈''
|label2=유키노돈트
|2={{clade
|1={{clade
|1=''키노그나투스''
|2={{clade
|1=트라베르소돈트과
|2=트리티로돈트과
}}
}}
|label2=프로베이노그나티아
|2={{clade
|1={{clade
|1=키니쿠돈트과
|2=트리텔로돈트과
}}
|2={{clade
|1=''프로조스트로돈''
|label2=포유류형류
|2={{clade
|1=모르가누코돈과
|2={{clade
|1=도코돈타
|2={{clade
|1=''하드로코디움''
|2={{clade
|1=쿠네오테리움과
|2=크라운 그룹(crown group) 포유류
}}
}}
}}
}}
}}
}}
}}
}}
}}

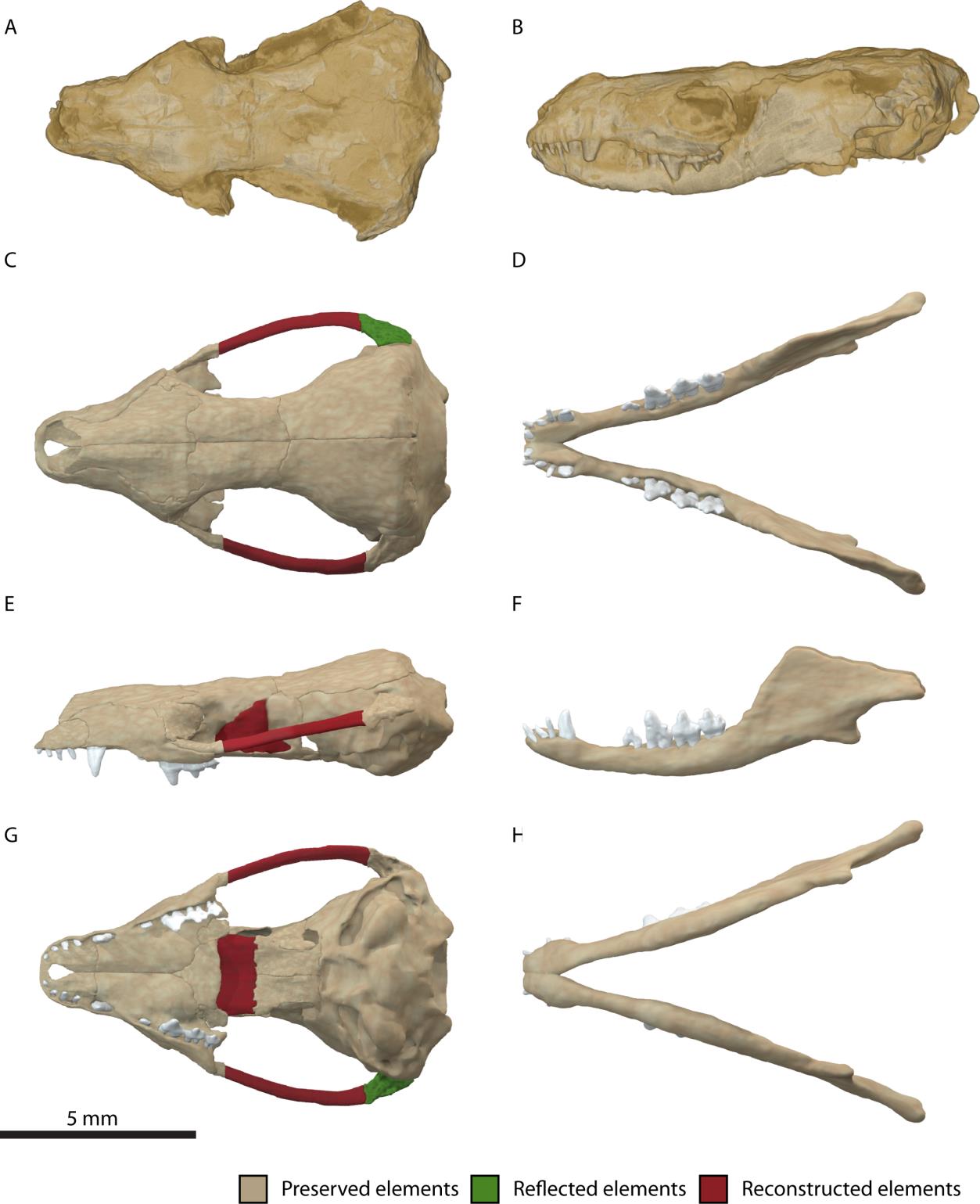

포유류 청각 소골 진화
초기 쥐라기 시대 화석이 발견된 ''하드로코디움''은 포유류의 턱 관절과 중이에 대한 최초의 명확한 증거를 제공한다. 턱 관절은 치골과 인두골로 형성되고, 관절골과 사각골은 중이로 이동하여 모루뼈와 망치뼈로 알려지게 된다.
2. 1. 단궁류의 출현
현존하는 포유류는 암컷의 젖샘으로 식별할 수 있지만, 화석에서는 젖샘과 같은 연조직이 보이지 않기 때문에 다른 특징을 사용해야 한다. 고생물학에서 사용하는 특징 중 하나는 현존하는 모든 포유류(단공류 포함)가 공유하지만 초기 트라이아스기 수궁류에서는 나타나지 않는 턱 관절 구조이다. 포유류는 치골(이를 지닌 아래턱 뼈)과 인상골(또 다른 작은 두개골 뼈)로만 구성된 턱 관절을 가지는 반면, 다른 모든 양막류는 관절뼈(아래턱 뒤쪽의 작은 뼈)와 네모뼈(위턱 뒤쪽의 작은 뼈)로 구성된 턱 관절을 가진다. 포유류의 턱 관절 구조는 쥐라기 시대에 네모뼈와 관절뼈가 중이의 침골과 망치뼈로 진화하면서 형성되었다.[5][6] 포유류는 또한 이중 후두 융기를 가지는데, 다른 사지동물은 단일 후두 융기를 가지는 반면, 포유류는 목의 최상위 척추뼈에 맞는 두개골 기저부에 두 개의 융기를 가지고 있다.
1981년 케네스 A. 커맥과 그의 동료들은 포유류와 초기 시냅시드 사이의 경계선을 포유류형 어금니 교합 패턴이 획득되고 치골-인상골 관절이 나타나는 지점으로 긋는 것을 제안했다. 이 기준은 "아래턱이 중생대 포유류의 보존될 가능성이 가장 높은 골격 요소"라는 사실에 근거한 편의상의 문제였다.[7] 오늘날 대부분의 고생물학자들은 이 기준을 충족하는 동물을 포유류로 간주한다.[39]
최초의 완전 육상 척추동물은 파충류 양막류였다. 이들의 알은 내부 막을 가지고 있어서 배아가 호흡하면서도 수분을 유지할 수 있었다. 이는 양막류가 건조한 육지에 알을 낳을 수 있게 해주었으며, 양서류는 일반적으로 물속에 알을 낳아야 했다. ( 수리남두꺼비와 같은 일부 양서류는 이러한 제약을 극복하기 위해 다른 방법을 개발했다). 최초의 양막류는 석탄기 중기에 조상인 파충형류로부터 나타났다.[8]
수백만 년 이내에 두 개의 중요한 양막류 계통이 분화되었는데, 단궁류는 포유류의 조상이며, 파충류는 도마뱀, 뱀, 거북, 악어, 공룡, 그리고 조류의 조상이다.[3] 단궁류와 파충류의 가장 초기의 화석(각각 ''아르케오티리스''와 ''힐로노무스'' 등)은 약 ~ 전으로 거슬러 올라간다. 척추동물 화석이 석탄기 후반에는 매우 희귀하기 때문에, 기원 시기를 정확히 알기는 어렵다. 따라서 각 동물 유형의 실제 최초 출현 시기는 최초의 화석 발견보다 훨씬 이전일 수 있다.[9]
2. 2. 수궁류의 번성
고생물학에서 현존하는 포유류는 암컷의 젖샘으로 식별할 수 있지만, 화석에서는 젖샘과 같은 연조직이 보이지 않기 때문에 다른 특징을 사용해야 한다. 모든 현존하는 포유류(단공류 포함)에게는 있지만 초기 트라이아스기 수궁류에게는 없는 특징은 다음과 같다. 포유류는 다른 양막류가 먹는 데 사용하는 두 개의 작은뼈를 청각에 사용한다. 초기 양막류는 관절뼈(아래턱 뒤쪽의 작은 뼈)와 네모뼈(위턱 뒤쪽의 작은 뼈)로 구성된 턱 관절을 가졌다. 양서류, 거북이, 도마뱀, 뱀, 악어류, 공룡(조류 포함), 어룡, 익룡 및 수궁류를 포함한 모든 비포유류 사지동물이 이 시스템을 사용한다. 그러나 포유류는 치골(이를 지닌 아래턱 뼈)과 인상골(또 다른 작은 두개골 뼈)로만 구성된 다른 턱 관절을 가지고 있다. 쥐라기 시대에 네모뼈와 관절뼈는 중이의 침골과 망치뼈로 진화했다.[5][6] 포유류는 또한 이중 후두 융기를 가지고 있는데, 다른 사지동물은 단일 후두 융기를 가지고 있는 반면, 목의 최상위 척추뼈에 맞는 두개골 기저부에 두 개의 융기를 가지고 있다.테라프시드는 초기 단궁류인 스페나코돈티아에서 과달루페 절 중기에 기원했으며, 육상 척추동물로서 그들을 대체하여 우위를 점했다. 그들은 더 큰 측두창과 크기가 같은 앞니를 포함하여 두개골과 턱의 여러 특징에서 이전의 단궁류와 다르다.[13]
테라프시드 계통은 여러 단계를 거쳐 페름기 후기에 키노돈의 진화를 이끌었고, 이 중 일부는 초기 포유류와 비슷해지기 시작했다:[14]
- 점진적인 뼈로 된 이차 경구개의 발달.
- 치골이 점차 아래턱의 주요 뼈가 되었다.
디노케팔리아(Dinocephalia, "끔찍한 머리")는 육식동물과 초식동물을 모두 포함했다. 그들은 컸는데, 예를 들어 ''안테오사우루스''(Anteosaurus)는 길이가 약 6.10m에 달했다. 육식동물 중 일부는 반-직립형 뒷다리를 가지고 있었지만, 모든 디노케팔리아는 옆으로 뻗은 앞다리를 가지고 있었다. 여러 면에서 그들은 매우 원시적인 수궁류였는데, 예를 들어 이차 구개가 없고 턱이 다소 "파충류" 같았다.[17]
이치류(이빨 이상 동물) ("이례적인 이빨")는 가장 성공적인 초식 수궁류 중 하나였으며, 그중 하위 그룹인 디키노돈트는 트라이아스기 말까지 살아남았다. 그러나 디키노돈트는 현대 초식 포유류와 매우 달랐는데, 이들의 유일한 이빨은 위턱에 있는 한 쌍의 송곳니였고 (몇몇 파생된 칸네마이에리iforms에서는 사라짐) 일반적으로 새나 각룡류와 같은 부리를 가지고 있었다고 여겨진다.[18]
수수류(Theriodonts, "짐승 이빨")와 그들의 후손들은 아래턱의 관절골이 두개골의 매우 작은 사각골을 단단히 움켜쥐는 턱 관절을 가지고 있었다. 이것은 훨씬 더 넓은 입을 벌릴 수 있게 했고, 육식성 고르고놉스("고르곤 얼굴") 그룹이 "검치"를 발달시킬 수 있게 했다. 그러나 수수류의 턱 경첩은 더 장기적인 중요성을 지녔다. 사각골의 크기가 매우 작아진 것은 포유류 턱 관절과 중이의 발달에 중요한 단계였다.
고르고놉스는 여전히 몇 가지 원시적인 특징을 가지고 있었다. 골질의 이차적 구개(올바른 위치의 다른 뼈가 동일한 기능을 수행함) 부재, 팔다리가 옆으로 뻗어 있음, 뻗은 자세와 직립 자세 모두에서 작동할 수 있는 뒷다리가 그것이다. 고르고놉스류와 거의 같은 시기에 나타난 것으로 보이는 테로케팔리아("짐승 머리")는 추가적인 포유류형 특징을 가지고 있었다. 예를 들어, 손가락과 발가락 뼈는 초기 포유류(그리고 영장류가 가지고 있는 것과 동일한 수의, 인간 포함)와 같은 수의 지골(분절)을 가지고 있었다.[19] 테로케팔리아와 불확실한 기저 시조류(전악류)에 속할 수 있는 많은 창싱기 분변화석에는 털의 잔해일 수 있는 길쭉한 속이 빈 구조가 포함되어 있다. 이는 테라푸시드가 2억 5200만 년 전부터 털로 덮여 있었음을 의미한다.[20]
추가 정보: 야행성 병목 현상
키노돈트(Cynodontia)는 후기 페름기에 등장한 수수류 그룹으로, 모든 포유류의 조상을 포함한다. 키노돈트의 포유류와 유사한 특징으로는 아래턱 뼈의 수 감소, 이차적인 경구개, 치관에 복잡한 패턴이 있는 볼 이빨, 그리고 두개강을 채우는 뇌가 있다.[21]
다중 챔버 굴이 발견되었는데, 초기 트라이아스기 키노돈 ''Trirachodon''의 20개 뼈가 들어 있었다. 이 동물들은 급작스런 홍수로 익사한 것으로 생각된다. 광범위한 공유 굴은 이 동물들이 복잡한 사회적 행동을 할 수 있었음을 나타낸다.[22]
이들의 원시적 단궁류와 수궁류 조상은 매우 컸지만(약 약 1.52m에서 약 2.44m), 키노돈트는 페름기-트라이아스기 대멸종 이전에도 다른 수궁류와의 경쟁 때문에 점차 크기가 감소했다(약 약 0.46m에서 약 1.52m). 대멸종 이후, 프로보노그나티아 키노돈트 그룹은 시조류와의 새로운 경쟁으로 인해 크기가 급격히 감소했고(약 약 10.16cm에서 약 45.72cm), 야행성으로 전환하여 야행성 특징, 폐포, 세기관지, 더 큰 호흡 표면적을 위한 발달된 횡격막, 핵이 없는 적혈구, 맹장 이후 진정한 결장이 있는 대장, 항온성, 털이 많고 분비샘이 있으며 체온 조절 기능이 있는 피부(피지와 땀을 분비), 그리고 높은 신진대사, 더 큰 뇌, 완전히 직립한 뒷다리를 유지하기 위한 4개의 챔버 심장을 진화시켰다(앞다리는 반쯤 엎드린 상태로 유지되었고, 나중에 수아목류에서만 그렇게 되었다). 일부 피부샘은 새끼의 신진대사 요구를 충족시키기 위해 암컷에서 유선으로 진화했을 수 있다(10배 증가). 많은 골격 변화도 일어났다. 예를 들어, 치골이 더 강해지고 분화된 치아를 유지했으며, 두개골의 코 구멍 쌍이 융합되었다.
이러한 진화적 변화는 최초의 포유류(크기 약 약 10.16cm)로 이어졌다. 그들은 빠른 성장과 짧은 수명을 진화시킨 것으로 보이며, 이러한 생활사는 수많은 현대 소형 포유류에서도 발견된다.[23] 그들은 또한 굴을 파는 생활 방식에 적응하여, 공룡이 이족 보행을 할 수 있게 해준 큰 꼬리 기반 다리 근육을 잃었는데, 이는 이족 보행 포유류가 매우 드문 이유를 설명할 수 있다.[24]
2. 3. 키노돈트에서 포유류로
테랍시드는 초기 단궁류인 스페나코돈티아에서 과달루페 절 중기에 기원했으며, 육상 척추동물로서 그들을 대체하여 우위를 점했다. 그들은 더 큰 측두창과 크기가 같은 앞니를 포함하여 두개골과 턱의 여러 특징에서 이전의 단궁류와 다르다.[13]테랍시드 계통은 여러 단계를 거쳐 페름기 후기에 키노돈의 진화를 이끌었고, 이 중 일부는 초기 포유류와 비슷해지기 시작했다:[14]
- 점진적인 뼈로 된 이차 경구개의 발달. 대부분의 책과 논문은 이것을 포유류의 높은 신진대사율 진화의 전제 조건으로 해석하는데, 이는 이러한 동물들이 동시에 먹고 숨을 쉴 수 있게 해주었기 때문이다. 그러나 일부 과학자들은 일부 현대 변온 동물들이 입과 기도를 분리하기 위해 살로 된 이차 구개를 사용하며, ''뼈로 된'' 구개는 혀가 음식을 조작하여 호흡보다는 씹는 것을 용이하게 하는 표면을 제공한다고 지적한다.[15] 뼈로 된 이차 구개를 씹는 데 도움이 된다고 해석하는 것은 또한 더 빠른 신진대사의 발달을 시사하는데, 씹는 것은 위로 전달되는 음식 입자의 크기를 줄이고 따라서 소화를 가속화할 수 있기 때문이다. 포유류에서 구개는 두 개의 특정 뼈로 형성되지만, 다양한 페름기 테랍시드는 구개 역할을 하기에 적절한 위치에 다른 뼈의 조합을 가지고 있었다.
- 치골이 점차 아래턱의 주요 뼈가 되었다.
다음은 포유류 진화와 관련된 주요 내용만 표시한 단순화된 계통수이다.[13]
{{clade| style=font-size:90%;line-height:100%
|label1=수궁류(Therapsids)
|1={{clade
|1=쌍궁류(Biarmosuchia)

|label2=진수궁류(Eutherapsida)
|2={{clade
|1={{clade
|1={{clade
|1=공룡두류(Dinocephalia)
|label2=신수궁류(Neotherapsida)
|2={{clade
|label1=이치류(Anomodont)
|1={{clade
|1=이궁류(Dicynodont)
}}
|label2=수수류(Theriodontia)
|2={{clade
|1=고르고놉스류(Gorgonopsia)
|label2=진수수류(Eutheriodontia)
|2={{clade
|1=수두류(Therocephalia)
|label2=개이빨류(Cynodontia)
|2={{clade
|1=(결국 포유류) --
}}
}}
}}
}}
}}
}}
}}
}}
트라이아스기까지 살아남은 것은 이궁류, 수두류, 개이빨류뿐이었다.
이치류(이빨 이상 동물) ("이례적인 이빨")는 가장 성공적인 초식 수궁류 중 하나였으며, 그중 하위 그룹인 디키노돈트는 트라이아스기 말까지 살아남았다. 그러나 디키노돈트는 현대 초식 포유류와 매우 달랐는데, 이들의 유일한 이빨은 위턱에 있는 한 쌍의 송곳니였고 (몇몇 파생된 칸네마이에리iforms에서는 사라짐) 일반적으로 새나 각룡류와 같은 부리를 가지고 있었다고 여겨진다.[18]
수수류(Theriodonts, "짐승 이빨")와 그들의 후손들은 아래턱의 관절골이 두개골의 매우 작은 사각골을 단단히 움켜쥐는 턱 관절을 가지고 있었다. 이것은 훨씬 더 넓은 입을 벌릴 수 있게 했고, 육식성 고르고놉스("고르곤 얼굴") 그룹이 "검치"를 발달시킬 수 있게 했다. 그러나 수수류의 턱 경첩은 더 장기적인 중요성을 지녔다. 사각골의 크기가 매우 작아진 것은 포유류 턱 관절과 중이의 발달에 중요한 단계였다.
테로케팔리아("짐승 머리")는 추가적인 포유류형 특징을 가지고 있었다. 예를 들어, 손가락과 발가락 뼈는 초기 포유류(그리고 영장류가 가지고 있는 것과 동일한 수의, 인간 포함)와 같은 수의 지골(분절)을 가지고 있었다.[19] 테로케팔리아와 불확실한 기저 시조류(전악류)에 속할 수 있는 많은 창싱기 분변화석에는 털의 잔해일 수 있는 길쭉한 속이 빈 구조가 포함되어 있다. 이는 테라푸시드가 2억 5200만 년 전부터 털로 덮여 있었음을 의미한다.[20]
추가 정보: 야행성 병목 현상
키노돈트(Cynodontia)는 후기 페름기에 등장한 수수류 그룹으로, 모든 포유류의 조상을 포함한다. 키노돈트의 포유류와 유사한 특징으로는 아래턱 뼈의 수 감소, 이차적인 경구개, 치관에 복잡한 패턴이 있는 볼 이빨, 그리고 두개강을 채우는 뇌가 있다.[21]
다중 챔버 굴이 발견되었는데, 초기 트라이아스기 키노돈 ''Trirachodon''의 20개 뼈가 들어 있었다. 이 동물들은 급작스런 홍수로 익사한 것으로 생각된다. 광범위한 공유 굴은 이 동물들이 복잡한 사회적 행동을 할 수 있었음을 나타낸다.[22]
이들의 원시적 단궁류와 수궁류 조상은 매우 컸지만(약 약 1.52m에서 약 2.44m), 키노돈트는 페름기-트라이아스기 대멸종 이전에도 다른 수궁류와의 경쟁 때문에 점차 크기가 감소했다(약 약 0.46m에서 약 1.52m). 대멸종 이후, 프로보노그나티아 키노돈트 그룹은 시조류와의 새로운 경쟁으로 인해 크기가 급격히 감소했고(약 약 10.16cm에서 약 45.72cm), 야행성으로 전환하여 야행성 특징, 폐포, 세기관지, 더 큰 호흡 표면적을 위한 발달된 횡격막, 핵이 없는 적혈구, 맹장 이후 진정한 결장이 있는 대장, 항온성, 털이 많고 분비샘이 있으며 체온 조절 기능이 있는 피부(피지와 땀을 분비), 그리고 높은 신진대사, 더 큰 뇌, 완전히 직립한 뒷다리를 유지하기 위한 4개의 챔버 심장을 진화시켰다(앞다리는 반쯤 엎드린 상태로 유지되었고, 나중에 수아목류에서만 그렇게 되었다). 일부 피부샘은 새끼의 신진대사 요구를 충족시키기 위해 암컷에서 유선으로 진화했을 수 있다(10배 증가). 많은 골격 변화도 일어났다. 예를 들어, 치골이 더 강해지고 분화된 치아를 유지했으며, 두개골의 코 구멍 쌍이 융합되었다.
이러한 진화적 변화는 최초의 포유류(크기 약 약 10.16cm)로 이어졌다. 그들은 빠른 성장과 짧은 수명을 진화시킨 것으로 보이며, 이러한 생활사는 수많은 현대 소형 포유류에서도 발견된다.[23] 그들은 또한 굴을 파는 생활 방식에 적응하여, 공룡이 이족 보행을 할 수 있게 해준 큰 꼬리 기반 다리 근육을 잃었는데, 이는 이족 보행 포유류가 매우 드문 이유를 설명할 수 있다.[24]
(Cynodontia:Dendrogram – Palaeos를 기반으로 함[40])
{{clade| style=font-size:90%;line-height:100%
|label1=키노돈트
|1={{clade
|1={{clade
|1=''드비니아''
|2=프로키노수키드
}}
|label2=에피키노돈트
|2={{clade
|1=''트리낙소돈''
|label2=유키노돈트
|2={{clade
|1={{clade
|1=''키노그나투스''
|2={{clade
|1=트라베르소돈트과
|2=트리티로돈트과
}}
}}
|label2=프로베이노그나티아
|2={{clade
|1={{clade
|1=키니쿠돈트과
|2=트리텔로돈트과
}}
|2={{clade
|1=''프로조스트로돈''
|label2=포유류형류
|2={{clade
|1=모르가누코돈과
|2={{clade
|1=도코돈타
|2={{clade
|1=''하드로코디움''
|2={{clade
|1=쿠네오테리움과
|2=크라운 그룹(crown group) 포유류
}}
}}
}}
}}
}}
}}
}}
}}
}}
포유류 청각 소골의 진화
초기 쥐라기 시대 화석이 발견된 ''하드로코디움''은 포유류의 턱 관절과 중이에 대한 최초의 명확한 증거를 제공한다. 턱 관절은 치골과 인두골로 형성되고, 관절골과 사각골은 중이로 이동하여 모루뼈와 망치뼈로 알려지게 된다.
2. 4. 초기 포유류형 동물
고생물학에서 현생 포유류와 화석을 구분하는 특징 중 하나는 턱 관절의 구조이다. 포유류는 치골(이를 지닌 아래턱 뼈)과 인상골(또 다른 작은 두개골 뼈)로만 구성된 턱 관절을 가지고 있다. 반면, 초기 양막류는 관절뼈(아래턱 뒤쪽의 작은 뼈)와 네모뼈(위턱 뒤쪽의 작은 뼈)로 구성된 턱 관절을 가졌다. 포유류의 쥐라기 시대에 네모뼈와 관절뼈는 중이의 침골과 망치뼈로 진화했다.[5][6]1981년 케네스 A. 커맥과 동료들은 포유류와 초기 시냅시드 사이의 경계선을 포유류형 어금니 교합 패턴과 치골-인상골 관절의 출현으로 정의했다.[7] 오늘날 대부분의 고생물학자들은 이 기준을 따르는 동물을 포유류로 간주한다.[39]
중생대의 단궁류는 이빨뼈와 비늘뼈로 구성된 턱 관절을 가졌지만, 대부분 쥐보다 작아 화석으로 보존되기 어려웠다. 이들은 화석화에 불리한 환경에 서식했고, 섬세한 뼈는 쉽게 파괴되었으며, 작은 화석은 발견되기 어려웠다. 그러나 지난 수십 년 동안 중생대 포유류 화석이 획기적으로 증가했다. 1979년에는 116개 속만 알려졌지만, 2007년에는 약 310개 속으로 증가했으며, 최소 18종의 중생대 포유류는 거의 완전한 골격으로 대표된다.[36]
모르가누코돈류는 약 2억 500만 년 전 트라이아스기 후기에 처음 등장했으며, 덴타리-스쿠아모살과 관절-사각 턱 관절을 모두 가지고 있어 과도기적 화석의 좋은 예시이다.[41]
도코돈트(Docodonts)는 가장 흔한 쥐라기 포유류형 동물 중 하나로, 정교한 어금니로 유명하다. 약 1억 6400만 년 전 쥐라기 중기에 살았던 어식성 동물인 ''카스토로카우다(Castorocauda)''("비버 꼬리")는 2004년에 처음 발견되어 2006년에 기술되었다. ''카스토로카우다''는 왕관군 포유류는 아니지만, 거의 완전한 골격으로 발견되었고, "작은 야행성 곤충 먹이"라는 고정관념을 깨뜨렸기 때문에 포유류 진화 연구에 매우 중요하다:[42]
- 코에서 꼬리 끝까지 약 약 43.18cm이고, 꼬리는 약 12.70cm이며, 무게는 500g에서 800g 정도였을 수 있다.
- 가장 오래된 확실한 털의 증거를 제공한다.
- 납작한 꼬리뼈와 뒷발가락 사이에 부드러운 조직의 잔재가 있어 물갈퀴가 있었음을 시사한다.
- ''카스토로카우다''의 강력한 앞다리는 땅을 파는 데 적합해 보인다.
- 이빨은 물고기를 먹는 데 적합해 보인다.
''하드로코디움''은 약 1억 9500만 년 전인 초기 쥐라기에 살았으며, 턱 관절이 인설골과 치골로만 구성되어 있고, 고막이 두개골의 일부였음을 시사한다. 관절골과 네모뼈는 중이로 이동하여 망치뼈와 모루뼈가 되었을 것이다.[43] ''하드로코디움''의 뇌와 턱은 서로 묶여 있지 않아, 뇌는 확장되고 턱은 먹는 것에 특화된 방식으로 적응할 수 있었다.
쿠네오테리움과는 후기 트라이아스기와 초기 쥐라기에 알려져 있으며, 치아 구조를 기반으로 '시메트르돈타' 또는 '판토테리아'의 일부로 분류되었으나, 이후 관 그룹 포유류의 가장 가까운 친척 중 하나로 밝혀졌다. 치아 화석만 발견되어 계통 발생 연구에 거의 포함되지 않는다.[44]
다구치류(여러 개의 "결절"이 "어금니"에 있어서 이름이 붙여짐)는 "중생대의 설치류"라고 불리지만, 수렴 진화의 한 예이다. 이들은 약 1억 2천만 년 동안 존재했으며, 초기 올리고세에 멸종했다.
일부 학자들은 다구치류를 포유류의 관 그룹에서 제외하고, 모르가누코돈보다 현존하는 포유류와 더 멀리 관련이 있다고 주장한다. 다구치류는 턱 관절이 치골과 인설골로만 구성되어 있다는 점에서 포유류와 유사하지만, 사각골과 관절골은 중이의 일부이다. 이들의 이빨은 분화되어 있고, 맞물리며, 포유류와 같은 치아 융기를 가지고 있으며, 관골궁을 가지고 있으며, 골반의 구조는 현대의 유대류와 같이 작고 무력한 새끼를 낳았음을 시사한다. 반면에 다음과 같은 점에서 현대 포유류와 다르다.
- "어금니"는 두 개의 평행한 결절 열을 가지고 있다.
- 저작 작용이 다르다.
- 관골궁의 앞쪽 부분은 상악골(위턱뼈)로 구성된다.
- 인설골은 뇌두개골의 일부를 형성하지 않는다.
- 코는 페리코사우루스와 더 비슷해 보인다.
포유류 청각 소골 진화
초기 쥐라기 시대 화석이 발견된 ''하드로코디움''은 포유류의 턱 관절과 중이에 대한 최초의 명확한 증거를 제공한다. 턱 관절은 치골과 인두골로 형성되고, 관절골과 사각골은 중이로 이동하여 모루뼈와 망치뼈로 알려지게 된다.
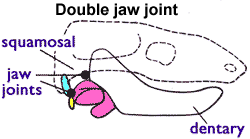
단공류 ''테이놀로포스''에 대한 한 분석에 따르면, 이 동물은 각골과 사각골로 형성된 포유류 이전의 턱 관절을 가지고 있었고, 결정적인 포유류의 중이는 단공류와 수아류 포유류에서 두 번 독립적으로 진화했다는 주장이 제기되었지만, 이 아이디어는 논쟁의 대상이 되었다.[116]
3. 현생 포유류의 분류

테리아(Theria, "짐승")는 진수류(Eutheria, 태반류 포함)와 후수류(Metatheria, 유대류 포함)의 마지막 공통 조상에서 유래된 분기도이다.
테리아의 공통적인 특징은 다음과 같다:[60]
- 쇄골간 부재.[56]
- 부리돌기가 존재하지 않거나, 견갑골과 융합되어 부리돌기를 형성.
- 족근골 발목 관절은 경골과 목말뼈 사이에 위치하며, 발꿈치뼈는 경골과 접촉하지 않고 근육이 부착될 수 있는 뒤꿈치를 형성한다.
- 삼첨형 어금니.[52]
크라운 그룹 포유류는 현존하는 포유류와 마지막 공통 조상까지의 가까운 친척을 포함한다. 이 그룹은 살아있는 구성원을 가지고 있기 때문에, DNA 분석을 통해 화석에서 나타나지 않는 특징의 진화를 설명하려는 시도가 이루어질 수 있다.
초기 양막류(Amniotes)는 망막의 원뿔 세포에 4개의 옵신을 가지고 있어 색상을 구별하는 데 사용했다.[46][47] 녹색 옵신은 어떤 관(crown) 포유류에게도 유전되지 않았지만, 모든 정상적인 개체는 빨간색 옵신을 물려받았다. 현존하는 모든 후손은 파란색에 민감한 옵신 중 하나를 잃었지만 항상 같은 것을 잃지는 않았다. 단공류는 파란색에 민감한 옵신 하나를 유지하고, 유대류와 태반류는 다른 하나를 유지한다(나중에 다른 파란색 옵신도 잃은 고래류(cetacea) 제외).[48] 영장류를 포함한 일부 태반류와 유대류는 이후 녹색에 민감한 옵신을 진화시켰다. 따라서 초기 관 포유류처럼 그들의 시력은 3색 시각(Trichromacy)이다.[49][50]
현생 포유류는 크게 단공류, 유대류, 진수류(태반류)로 나뉜다.
3. 1. 단공류
현존하는 포유류 중 가장 원시적인 것은 단공류이며, 모두 오스트레일리아의 열대권에서만 서식한다. 몸은 대개 털과 가시로 덮여 있는데, 그 중 오리너구리 같은 것은 주둥이가 각질이어서 오리와 비슷하다. 같은 단공류에는 가시두더지가 있다. 이들 동물은 땅 속에 구멍을 파고 들어가 사는데 모두가 난생이고 새끼를 기른다.오스트랄로스페니다는 오스트리보스페니데와 단공류를 포함하기 위해 정의된 그룹이다. ''아스팔토미로스''(쥐라기 중기~후기, 파타고니아)는 기초 오스트랄로스페니드로 해석되었으며, 오스트랄로스페니드가 곤드와나 (고대 남반구 초대륙) 전역에 널리 퍼져 있었음을 보여준다.[53]
1억 2100만 년에서 1억 1250만 년 사이에 살았던 ''테이놀로포스''에 대한 최근 분석에 따르면 이는 "크라운 그룹"(진화된, 상대적으로 특화된) 단공류였다. 이는 기초(가장 원시적인) 단공류가 훨씬 더 일찍 나타났음에 대한 증거로 받아들여졌지만, 이에 대한 논란이 있었다. 이 연구는 또한 일부 오스트랄로스페니드로 여겨졌던 동물들이 "크라운 그룹" 단공류(예: ''스테로포돈'')였고, 다른 오스트랄로스페니드로 여겨졌던 동물(예: ''오스트리보스페노스'', ''비숍스'', ''암본드로'', ''아스팔토미로스'')이 유대류와 태반류를 포함하는 그룹인 테리안 포유류와 더 밀접한 관련이 있으며, 아마도 그 구성원일 가능성이 있음을 시사했다.[54]
호주에서 발견된 ''테이놀로포스''(Teinolophos)는 가장 오래된 단공류로 알려져 있다. 2007년 연구(2008년 발표)에 따르면, 테이놀로포스는 기저 단공류가 아니라 완전한 오리너구리이며, 따라서 오리너구리와 바늘두더지 계통은 훨씬 이전에 분화되었다고 한다.[54] 하지만 최근 연구(2009년)에 따르면, ''테이놀로포스''는 일종의 오리너구리이면서도 기저 단공류였으며, 현생 단공류의 방산보다 앞섰다고 한다. 오리너구리의 반수생 생활 방식은 수백만 년 전에 호주로 이주한 유대류와의 경쟁을 피하게 해주었는데, 이는 새끼 주머니쥐가 어미에게 붙어 있어야 하고 어미가 물에 들어가면 익사하기 때문이다(하지만 수생 주머니쥐와 수달 주머니쥐와 같은 예외도 있다. 그러나 이들은 모두 남아메리카에 살며 단공류와 접촉하지 않는다). 유전적 증거에 따르면 바늘두더지는 반수생에서 육상 생활 방식으로 전환하면서 오리너구리 계통에서 1900만~4800만 년 전에 분화되었다.[55]
단공류는 키노돈트 조상으로부터 물려받았을 수 있는 몇 가지 특징을 가지고 있다.
- 도마뱀과 새와 마찬가지로, 소변, 배변 및 생식을 위해 동일한 구멍을 사용한다("단공류"는 "하나의 구멍"을 의미한다).
- 도마뱀, 거북 및 악어류와 같이 가죽 같고 석회화되지 않은 알을 낳는다.
- 다른 포유류와 달리, 암컷 단공류는 젖꼭지가 없으며 배의 반점에서 "땀"으로 우유를 분비하여 새끼에게 먹이를 준다.
이러한 특징은 화석에서는 보이지 않으며, 고생물학자의 관점에서 주요 특징은 다음과 같다.[51]
- 근상돌기가 작거나 없는 가늘어진 치골 뼈.
- 귀의 외부 개구부는 턱의 후방 기저부에 위치한다.
- 협골 뼈가 작거나 없다.
- 강력한 복부 요소인 부리돌기, 쇄골 및 쇄골간을 가진 원시적인 견갑대. 참고: 테리아 포유류는 쇄골간이 없다.[56]
- 펼쳐지거나 반쯤 펼쳐진 앞다리.
3. 2. 유대류
현생 포유류 중 유대류(有袋類)는 새끼를 빨리 낳아 어미의 보육낭 속에서 키운다. 새끼는 어미의 젖을 먹으며 일정 크기로 자랄 때까지 밖으로 나오지 않는다. 유대류 화석은 유럽과 북아메리카의 중생대 지층에서도 나타났지만, 현재는 남아메리카와 오스트레일리아에만 서식한다. 북아메리카에 사는 버지니아주머니쥐 등은 남아메리카에서 건너온 것이다. 유대류는 형태와 생태가 다양하여, 유태반류의 설치류(齧齒類), 식충류(食蟲類), 빈치류(貧齒類), 식육류(食肉類) 등과 비슷한 것이 있다.현존하는 Metatheria는 모두 유대류이다. 몽골 후기 백악기의 ''Asiatherium''과 같은 몇몇 화석 속은 유대류이거나 다른 메타테리아 그룹의 구성원일 수 있다.[61][62]
가장 오래된 메타테리아는 중국 북동부 랴오닝성의 1억 2,500만 년 전의 초기 백악기 셰일에서 발견된 ''Sinodelphys''이다. 화석은 거의 완전하며 털 뭉치와 연조직의 흔적이 포함되어 있다.[63]
Didelphimorphia (서반구의 흔한 주머니쥐)는 후기 백악기에 처음 등장했으며 여전히 살아있는 대표자를 가지고 있는데, 이는 아마도 대부분 반-수목 비특수 잡식성이기 때문일 것이다.[64]
유대류의 가장 잘 알려진 특징은 번식 방식이다.
- 어미는 배아에 영양분을 공급하는 일종의 난황 낭을 자궁에 발달시킨다. 반디쿠트, 코알라, 웜뱃의 배아는 또한 자궁벽에 연결되는 태반과 같은 기관을 형성하지만, 태반과 같은 기관은 태반 포유류보다 작으며 어미로부터 배아로 영양분을 전달하는지는 확실하지 않다.[66]
- 임신 기간은 매우 짧으며 일반적으로 4~5주이다. 배아는 발달의 매우 초기 단계에서 태어나며, 일반적으로 태어날 때 약 5.08cm보다 짧다. 짧은 임신 기간은 어미의 면역 체계가 배아를 공격할 위험을 줄이기 위해 필요하다는 주장이 제기되었다.
- 새로 태어난 유대류는 앞다리(상대적으로 강한 손)를 사용하여 일반적으로 어미의 배에 있는 주머니 안의 젖꼭지로 기어 올라간다. 아기는 젖을 빨 힘이 없으므로 어미는 유선 위 근육을 수축시켜 아기에게 먹이를 준다. 새로 태어난 유대류가 젖꼭지로 기어 올라가기 위해 앞다리를 사용해야 한다는 것은 역사적으로 메타테리아 진화를 제한하는 것으로 여겨졌는데, 앞다리가 날개, 발굽 또는 지느러미와 같은 구조로 전문화될 수 없다고 가정했기 때문이다. 그러나 몇몇 반디쿠트, 특히 돼지발반디쿠트는 태반 유제류와 유사한 진짜 발굽을 가지고 있으며, 몇몇 유대류 활강류가 진화했다.

몇몇 유대류는 일부 태반류와 매우 유사하게 보이지만(주머니고양이 "유대류 호랑이" 또는 "유대류 늑대"가 좋은 예시이다), 유대류 골격은 태반류와 구별되는 몇 가지 특징을 가지고 있다.[67]
- 일부(주머니고양이 포함)는 4개의 어금니를 가지고 있다. 반면 알려진 태반류는 3개 이상을 가지고 있지 않다.
- 모두 두개골 바닥에 한 쌍의 구개 창(작은 콧구멍과 함께)이 있다.
유대류는 또한 암컷의 주머니를 지지하는 한 쌍의 유대류 뼈(때로는 "에피푸빅 뼈"라고 함)를 가지고 있다. 그러나 이들은 유대류에만 고유한 것은 아닌데, 이 뼈가 다구치류, 단공류, 심지어 진수류의 화석에서도 발견되었기 때문이다. 따라서 이들은 살아있는 태반 포유류의 조상이 유대류와 갈라진 후 어느 시점에서 사라진 일반적인 조상 특징일 가능성이 높다.[68][69] 일부 연구자들은 에피푸빅 뼈의 원래 기능이 허벅지를 앞으로 당기는 근육을 지지하여 이동에 도움을 주는 것이라고 생각한다.[70]
3. 3. 유태반류
유태반류는 새끼가 어미의 태반 안에서 오랫동안 자라는 것이 특징이며, 가장 오래된 화석은 식충류이다. 이들은 백악기부터 서식했는데, 크기가 작고 나무 위에서 생활했다. 그 후 다양한 형태의 유태반류가 나타났는데, 현존하는 개미핥기, 나무늘보, 아르마딜로를 포함한 남아메리카의 빈치류와 아시아의 천산갑, 아프리카의 땅돼지 등 원시적인 형태를 가진 종류는 모두 열대나 아열대에 서식하고 있다. 이들의 화석은 주로 신생대의 지층에서 발견된다.[18]유태반류는 진화하면서 점차 몸집이 커졌고, 식성도 다양해져 식물을 먹는 초식동물과 동물의 고기를 먹는 육식동물이 생겨났다. 초식동물에는 장비목, 유제류와 같이 몸집이 크고 무리를 지어 생활하는 것이 출현했고, 육식동물로는 표범, 호랑이, 여우 등과 같이 무리를 짓지 않고 생활하는 것이 많다. 이리도 육식을 주로 하지만, 겨울철에 먹이가 적을 때는 무리를 지어 대형 유제류 등을 습격하기도 한다. 잡식성 동물로는 곰 등이 있다.[18]
가장 초기의 진수류(Eutheria)가 나타난 시기는 논쟁의 대상이다. ''유라마이아'' 화석은 1억 6천만 년 전으로 거슬러 올라가며 진수류로 분류되었고,[71] 초기 백악기 1억 2천 5백만 년 전의 ''에오마이아'' 화석 또한 진수류로 분류되었다.[72] 그러나 최근의 형질 분석에 따르면, ''에오마이아''는 진수류 이전으로 분류되었으며, 가장 초기의 명확한 진수류 표본은 9천 1백만 년 전으로 거슬러 올라가는 ''마엘레스테스''에서 나왔다고 보고되었다.[73]

''에오마이아''는 유대류 및 초기 후수류와 더 유사한 몇 가지 특징을 가지고 있다.
- 골반에서 앞으로 뻗어 있는 치골은 현대의 태반류에서는 발견되지 않지만, 초기 포유류형류, 비태반 진수류, 유대류, 단공류 등 다른 모든 포유류에서 발견된다.[74]
- 좁은 골반 출구는 새끼가 태어날 때 매우 작았고 임신 기간이 짧았다는 것을 나타내며, 이는 현대의 유대류와 같다. 이는 태반이 나중에 발달했음을 시사한다.[75]
- 위턱의 각 측면에 5개의 앞니를 가지는데, 이는 후수류의 전형적인 특징이다. 현대 태반류의 치열에서 최대 숫자는 동형치아인 아르마딜로를 제외하고 3개이다. 그러나 ''에오마이아''의 어금니와 소구치의 비율은 진수류의 전형적인 특징이다.[74]
- ''에오마이아''는 또한 현대 태반류에서는 발견되지 않는 하악의 원시적인 특징인 메켈 연골을 가지고 있다.
태반류의 가장 잘 알려진 특징은 번식 방법이다.
- 배아는 어미에게 음식과 산소를 공급하고 노폐물을 제거하는 거대한 태반을 통해 자궁에 부착된다.
- 임신 기간은 비교적 길고 새끼는 태어날 때 상당히 잘 발달된다. 일부 종에서는 새끼가 태어난 지 한 시간 이내에 걷고 심지어 달릴 수도 있다.
태반 번식의 진화는 레트로바이러스에 의해 가능해졌다는 주장이 제기되었다.[76][77]
- 태반과 자궁 사이의 경계를 합포체로 만들어 산소, 영양분, 노폐물의 통과는 허용하지만, 어미의 면역 체계가 태아를 공격하게 하는 혈액 및 기타 세포의 통과는 방지한다.
- 어미의 면역 체계의 공격성을 감소시켜 태아에게는 좋지만 어미를 감염에 더 취약하게 만든다.
분자 계통 발생학은 화석의 특징을 고생물학자들이 사용하는 것과 거의 같은 방식으로 생물의 유전자 특징을 사용하여 계통수를 연구한다. 분자 계통 발생학자들은 고생물학자들의 계통수와 대체로 유사하면서도 주목할 만한 차이점을 보이는 계통수를 제안했다.
분자 계통 발생학자들의 계통수와 고생물학자들의 계통수 사이의 주요 차이점은 다음과 같다.
- 최상위 분할은 대서양상목과 북방진수류 사이에 있다.
- 아프리카진수류는 고생물학자 버전에서는 멀리 관련된 여러 그룹을 포함한다.
- 멸종된 목인 곤충류의 구성원은 세 개의 분류군으로 나뉜다.
- 박쥐는 영장류와 날원숭이보다 식육목 및 기제목에 더 가깝다.
- 기제목(홀수 발굽을 가진 유제류)은 우제목(짝수 발굽을 가진 유제류)보다 식육목과 박쥐에 더 가깝다.
4. 포유류의 생태와 다양성
키노돈트(Cynodontia)는 후기 페름기에 등장한 수수류 그룹으로, 모든 포유류의 조상을 포함한다.[21] 초기 트라이아스기 키노돈 ''Trirachodon'' 화석은 다수의 개체가 함께 굴속에서 발견되었는데, 이는 이들이 복잡한 사회적 행동을 했을 가능성을 보여준다.[22]
수수류와 그 후손들은 넓은 입을 벌릴 수 있는 턱 관절을 가졌고, 육식성 고르고놉스 그룹은 "검치"를 발달시켰다.[19] 테로케팔리아는 초기 포유류와 같은 수의 손발가락 뼈를 가졌다.[20]
대부분의 포유류는 작고 야행성이었지만, 일부는 더 크고 다양한 생활 방식을 가졌다.
포유류의 소화계는 식성에 따라 다양하게 진화했다. 육식 동물은 단순한 큰창자를, 초식 동물은 부피가 큰 큰창자를 가졌다.[124] 반추동물은 여러 개의 방으로 구성된 위를 가졌다.[125] 위산은 장내 미생물 군집 형성에 중요한 역할을 했다.[126] 인간 위장관 미생물군은 포유류 종의 진화와 함께 다양해졌다.[129][130]
4. 1. 서식 환경
포유류는 중생대 말기에 등장했는데, 이 시대에는 현재의 캥거루와 같은 유대류가 모두 나무 위에서 생활하고 있었다. 백악기 말이 되면서 이들이 출현하게 되었고, 이 시대에 식충류도 출현하여 모두 수상생활을 하고 있었다. 그러다가 초원이 퍼져감에 따라 평지에서 사는 포유류가 증가했다.약 2억 5200만 년 전 페름기 말의 대멸종은 육상 척추동물 종의 약 70%와 대부분의 육상 식물을 죽였다.[25] 그 결과, 생태계와 먹이 사슬이 붕괴되었고, 새로운 안정적인 생태계가 구축되기까지 약 3천만 년이 걸렸다.
아르코사우루스류는 빠르게 지배적인 육식 동물이 되었으며,[26] 이러한 발전을 종종 "트라이아스기 점령"이라고 부른다. 초기 트라이아스기가 주로 건조했기 때문에 아르코사우루스류의 우수한 물 보존 능력이 결정적인 이점을 제공했기 때문일 수 있다. 알려진 모든 아르코사우루스류는 샘이 없는 피부를 가지고 있으며, 물이 거의 포함되지 않은 질소 폐기물을 요산 페이스트로 배출하는 반면, 키노돈트는 오늘날 포유류가 하는 것처럼 대부분의 폐기물을 요소 용액으로 배출했을 가능성이 높다. 요소가 용해되도록 유지하려면 상당한 양의 물이 필요하다.[27]
트라이아스기 점령은 포유류의 진화에 매우 중요한 요인이었을 것이다. 초기 키노돈트에서 파생된 두 그룹은 아르코사우루스류와의 경쟁이 최소화된 틈새 시장에서 성공했다. 즉, 트리티로돈트류는 초식동물이었고, 포유류는 대부분 작은 야행성 곤충 식성이었다. 그 결과 포유류 조상의 몸 길이가 약 10.16cm 이하로 떨어지면서, 단열 및 체온 조절의 발전은 야행성 생활에 필수적이 되었을 것이다.[30]
도코돈트(Docodonts)는 가장 흔한 쥐라기 포유류형 동물 중 하나로, 정교한 어금니로 유명하다. 이들은 전반적으로 반수생 경향을 보였던 것으로 생각되며, 약 1억 6400만 년 전 쥐라기 중기에 살았던 어식성 동물인 ''카스토로카우다(Castorocauda)''("비버 꼬리")가 가장 잘 알려진 예시이다.[42]
일반적으로 대부분의 포유류형은 작고 야행성인 곤충의 생태적 지위를 차지했지만, 최근 주로 중국에서 발견된 결과에 따르면 일부 종, 특히 관군 포유류는 더 크고 과거 생각보다 더 다양한 생활 방식을 가졌다는 것을 보여준다.
- 테리아인 ''Patagomaia''는 후기 백악기 Chorrillo Formation (아르헨티나)에서 발견되었으며, 가장 큰 중생대 포유류이다.[80]
- ''Adalatherium hui''는 마다가스카르 백악기에서 발견된 크고, 똑바로 선 팔다리를 가진 초식 동물이다.[81]
- ''Castorocauda''는 약 1억 6400만 년 전 중기 쥐라기에 살았던 도코돈타의 일원으로, 비버와 유사한 꼬리는 수영에 적응되었고, 팔다리는 수영과 땅파기에 적응되었으며, 이빨은 물고기를 먹는 데 적응되었다.[42]
- 다구치류는 1억 2500만 년 이상 (약 1억 6천만 년 전 중기 쥐라기부터 약 3500만 년 전 후기 시신세) 생존한 알로테리아이다.
- 약 1억 5천만 년 전 후기 쥐라기 시대의 ''Fruitafossor''는 다람쥐와 비슷한 크기였으며 이빨, 앞다리, 등은 사회성 곤충의 둥지를 부수고 포식하는 데 적합했다.[84]
- 약 1억 2500만 년 전인 초기 백악기 경계에 살았던 ''Volaticotherium''은 가장 초기에 알려진 활강 포유류이다.[85]
- 1억 3천만 년 전 초기 백악기의 에우트리코노돈트인 ''Repenomamus''는 튼튼하고 오소리와 같은 포식자로, 때때로 어린 공룡을 잡아먹었다.[87][88]
- Zhelestidae는 후기 백악기의 초식 진수류 계통이다.[89]
- Deltatheroida는 육식 습성에 특화된 metatheria이다.[90]
- 모로코의 Berriasian에서 발견된 에우트리코노돈트인 ''Ichthyoconodon''은 해양 포유류였으며, 중생대에서 알려진 몇 안 되는 예 중 하나였을 가능성이 있음을 의미한다.[91]
- ''Didelphodon''은 후기 백악기의 스테고돈트과 유대류형 종으로, 담수 게와 연체동물을 먹는 데 특화되었을 것이다.
- 초기 백악기 Calonda Formation (앙골라)에서 너구리 크기의 포유류형의 발자국이 Mateus ''et al.'' (2017)에 의해 묘사되었으며, 새로운 이크노탁손 ''Catocapes angolanus''를 명명했다.
- 고비코노돈트가 훨씬 더 큰 공룡을 공격하는 모습이 보존되었다.[92]
4. 2. 식성
유태반류는 새끼가 어미의 태반 안에서 오랫동안 자라는 동물로, 가장 오래된 화석은 식충류이다. 백악기부터 서식했는데, 뒤쥐와 같이 작았으며 나무 위에서 생활했다. 그 후 여러 가지 형태의 유태반류가 출현했는데, 현존하는 것 중에서 개미핥기, 나무늘보, 아르마딜로를 포함한 남아메리카의 빈치류와 아시아의 천산갑, 아프리카의 땅돼지 등 원시적인 형태를 지닌 종류는 모두 열대나 아열대에 서식하고 있다. 이들의 화석은 신생대의 것이 주로 발견되고 있다. 유태반류는 진화하면서 점차 몸집이 커졌고, 식성도 양분되어 식물을 먹는 초식동물과 동물의 고기를 먹이로 먹는 육식동물이 생겨났다. 초식동물에는 장비류, 유제류와 같이 몸집이 크고 무리를 지어 생활하는 것이 출현했고, 육식동물로는 표범, 호랑이, 여우 등과 같이 무리를 짓지 않고 생활하는 것이 많다. 이리도 육식을 주로 하지만, 겨울철에 먹이가 적을 때는 무리를 지어 대형 유제류 등을 습격하기도 한다. 잡식성 동물로는 곰 등이 있다.[19]수수류와 그들의 후손들은 아래턱의 관절골이 두개골의 매우 작은 사각골을 단단히 움켜쥐는 턱 관절을 가지고 있었다. 이것은 훨씬 더 넓은 입을 벌릴 수 있게 했고, 육식성 고르고놉스 그룹이 "검치"를 발달시킬 수 있게 했다.
테로케팔리아는 손가락과 발가락 뼈는 초기 포유류와 같은 수의 지골(분절)을 가지고 있었다.[20]
일반적으로 대부분의 포유류형은 작고 야행성인 곤충의 생태적 지위를 차지했지만, 일부 종, 특히 관군 포유류는 더 크고 과거 생각보다 더 다양한 생활 방식을 가졌다는 것을 보여준다.
- 테리아인 ''Patagomaia''는 가장 큰 중생대 포유류이다.[80]
- ''Adalatherium hui''는 크고, 똑바로 선 팔다리를 가진 초식 동물이다.[81]
- ''Castorocauda''는 약 1억 6400만 년 전 중기 쥐라기에 살았던 도코돈타의 일원으로, 비버와 유사한 꼬리는 수영에 적응되었고, 팔다리는 수영과 땅파기에 적응되었으며, 이빨은 물고기를 먹는 데 적응되었다.[42]
- 다구치류는 1억 2500만 년 이상 생존한 알로테리아로 종종 "중생대의 설치류"라고 불린다.
- 약 1억 5천만 년 전 후기 쥐라기 시대의 ''Fruitafossor''는 다람쥐와 비슷한 크기였으며 이빨, 앞다리, 등은 사회성 곤충의 둥지를 부수고 포식하는 데 적합했다.[84]
- 고비코노돈트과의 ''Spinolestes''는 땅을 파는 데 적응되었고, 개미핥기류와 같은 습성을 가졌을 수도 있다.
- 약 1억 2500만 년 전인 초기 백악기 경계에 살았던 ''Volaticotherium''은 가장 초기에 알려진 활강 포유류로, 현대의 날다람쥐와 유사하게 팔다리 사이에 뻗어 있는 활강 막을 가지고 있었다.[85]
- 1억 3천만 년 전 초기 백악기의 에우트리코노돈트인 ''Repenomamus''는 튼튼하고 오소리와 같은 포식자로, 때때로 어린 공룡을 잡아먹었다. 두 종이 확인되었으며, 하나는 길이가 1m 이상이고 무게가 약 12kg에서 14kg이며, 다른 하나는 길이가 0.5m 미만이고 무게가 4kg에서 6kg이다.[87][88]
- ''Schowalteria''는 현대의 유제류와 유사하게 초식성으로의 분화를 보인다.
- Zhelestidae는 후기 백악기의 초식 진수류 계통이다.[89]
- mesungulatid도 후기 백악기의 비교적 큰 초식 포유류이다.
- Deltatheroida는 육식 습성에 특화된 metatheria이다.[90]
- 모로코의 Berriasian에서 발견된 에우트리코노돈트인 ''Ichthyoconodon''은 해양 포유류였으며, 중생대에서 알려진 몇 안 되는 예 중 하나였을 가능성이 있음을 의미한다.[91]
- ''Didelphodon''은 후기 백악기의 스테고돈트과 유대류형 종으로, 경골성 치아, 현대의 태즈메이니아 데빌과 유사한 튼튼한 턱을 가지고 있다. 담수 게와 연체동물을 먹는 데 특화되었을 것이다.
- 초기 백악기 Calonda Formation에서 너구리 크기의 포유류형의 발자국이 발견되었으며, ''Catocapes angolanus''를 명명했다.
- 고비코노돈트가 훨씬 더 큰 공룡을 공격하는 모습이 보존되었다.[92]
포유류의 소화계 진화는 포유류 진화에 상당한 영향을 미쳤다. 포유류의 출현과 함께 소화계는 동물의 식단에 따라 다양한 방식으로 수정되었다. 예를 들어, 고양이와 대부분의 육식 동물은 단순한 큰창자를 가지고 있는 반면, 초식 동물인 말은 부피가 큰 큰창자를 가지고 있다.[124] 반추동물의 조상 특징은 약 5천만 년 전에 진화한 여러 개의 방(보통 4개의 방)으로 구성된 위를 가지고 있다는 것이다.[125] 위산은 척추동물 장에서 발견되는 미생물 군집의 다양성과 구성을 형성하는 핵심 요소로 제안되었다. 포유류와 조류 분류군에서 영양 단계에 따른 위산도 비교 결과, 청소동물과 육식동물은 초식동물 또는 곤충이나 물고기와 같이 계통 발생학적으로 먼 먹이를 먹는 육식동물에 비해 위산도가 훨씬 더 높은 것으로 나타났다.[126]
인간 위장관 미생물군은 포유류 종이 진화함에 따라 함께 다양해졌다. 최근 연구에 따르면 포유류 종 간의 적응적 분화는 부분적으로 장내 미생물 군집의 변화에 의해 형성된다.[129][130]
4. 3. 사회 구조
키노돈트(Cynodontia)는 후기 페름기에 등장한 수수류 그룹으로, 모든 포유류의 조상을 포함한다. 키노돈트의 포유류와 유사한 특징으로는 아래턱 뼈의 수 감소, 이차적인 경구개, 치관에 복잡한 패턴이 있는 볼 이빨, 그리고 두개강을 채우는 뇌가 있다.[21]초기 트라이아스기 키노돈 ''Trirachodon''의 20개 뼈가 들어 있는 다중 챔버 굴이 발견되었다. 이 동물들은 급작스런 홍수로 익사한 것으로 생각된다. 광범위한 공유 굴은 이 동물들이 복잡한 사회적 행동을 할 수 있었음을 나타낸다.[22]
5. 포유류와 인간
(소스에 내용 없음)
참조
[1]
논문
New Jurassic Mammals from Patagonia, Argentina: A Reappraisal of Australosphenidan Morphology and Interrelationships
https://www.biodiver[...]
2007
[2]
웹사이트
Introduction to the Synapsida
http://www.ucmp.berk[...]
University of California Museum of Paleontology
1997-02-02
[3]
웹사이트
Amniota – Palaeos
http://palaeos.com/V[...]
2005-05-18
[4]
논문
Large Mesozoic mammals fed on young dinosaurs
http://doc.rero.ch/r[...]
2005-01-13
[5]
웹사이트
Mammalia: Overview – Palaeos
http://www.palaeos.c[...]
[6]
서적
History of Life
Blackwell Science
[7]
논문
The skull of ''Morganucodon''
1981-01
[8]
문서
Carroll R.L. (1991): The origin of reptiles. In: Schultze H.-P., Trueb L., (ed) ''Origins of the higher groups of tetrapods — controversy and consensus''. Ithaca: Cornell University Press, pp 331–353.
[9]
웹사이트
Synapsida: Varanopseidae – Palaeos
http://palaeos.com/v[...]
[10]
웹사이트
The Stem-Mammals—a Brief Primer
https://blogs.scient[...]
[11]
논문
Dimetrodon is Not a Dinosaur: Using Tree Thinking to Understand the Ancient Relatives of Mammals and their Evolution
[12]
뉴스
Meet the Ancient Reptile that Gave Rise to Mammals
https://www.scientif[...]
2016
[13]
웹사이트
Therapsida
http://www.palaeos.c[...]
[14]
서적
The evolution of mammalian characters
Croom Helm
[15]
서적
The Ecology and biology of mammal-like reptiles
Smithsonian Institution Press
1986
[16]
웹사이트
Therapsida: Biarmosuchia
http://palaeos.com/v[...]
[17]
웹사이트
Therapsida: Dinocephalia
http://palaeos.com/v[...]
[18]
웹사이트
Ammodontia
http://palaeos.com/v[...]
[19]
웹사이트
Theriodontia – Paleos
http://palaeos.com/v[...]
[20]
서적
Древнейшая находка остатков волос в копролитах тетрапод из терминальной перми Владимирской области
http://www.ginras.ru[...]
Paleontological Institute, Russian Academy of Sciences
[21]
웹사이트
Cynodontia Overview
http://palaeos.com/v[...]
[22]
논문
Vertebrate Burrow Complexes from the Early Triassic Cynognathus Zone (Driekoppen Formation, Beaufort Group) of the Karoo Basin, South Africa
2001-04-01
[23]
논문
The origin of placental mammal life histories
2022-10-06
[24]
웹사이트
New paper explains why predatory dinosaurs walked on two feet while mammals stayed on all fours
https://www.zmescien[...]
2017-03-03
[25]
웹사이트
Olenekian Age of the Triassic – Palaeos
http://www.palaeos.c[...]
[26]
서적
Vertebrate Palaeontology
Blackwell Science
[27]
서적
Comparative Animal Physiology
W. B. Sauders
[28]
문서
Darren Naish, Episode 38: A Not Too Shabby Podcarts
http://tetzoo.com/po[...]
[29]
논문
Trucidocynodon riograndensis gen. nov. et sp. nov. (Eucynodontia), a new cynodont from the Brazilian Upper Triassic (Santa Maria Formation)
[30]
논문
Selective Factors Associated with the Origin of Fur and Feathers
2000-08
[31]
논문
Fossil evidence on origin of the mammalian brain
2011-05
[32]
논문
Appraising the brain's energy budget
2002-08
[33]
간행물
Brain power
https://www.newscien[...]
2006-09-13
[34]
논문
Evolution of colour vision: The story of lost visual pigments
http://www.perceptio[...]
[35]
논문
The nocturnal bottleneck and the evolution of activity patterns in mammals
2013-08
[36]
논문
Transformation and diversification in early mammal evolution
2007-12
[37]
논문
Definition, diagnosis, and origin of Mammalia
1988-09-23
[38]
웹사이트
"Microlestes rhaeticus'' Dawkins 1864 (mammal)"
https://paleobiodb.o[...]
2012-01-30
[39]
서적
The Origin and Evolution of Mammals
Oxford University Press
[40]
웹사이트
Palaeos Vertebrates Cynodontia: Dendrogram
http://palaeos.com/v[...]
[41]
웹사이트
Morganucodontids & Docodonts – Palaeos
http://www.palaeos.c[...]
[42]
논문
A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals
http://doc.rero.ch/r[...]
2006-02
[43]
논문
A New Mammaliaform from the Early Jurassic and Evolution of Mammalian Characteristics
2001-05-25
[44]
논문
The Kuehneotheriidae (Mammaliaformes) from Saint-Nicolas-de-Port (Upper Triassic, France): a Systematic Review
https://link.springe[...]
2016
[45]
서적
Mammals of Europe: past, present, and future
Springer
2020
[46]
논문
Evolution of colour vision in mammals
2009-10-12
[47]
논문
Visual pigments in a living fossil, the Australian lungfish Neoceratodus forsteri
2007
[48]
논문
Evolution and Ecology of Retinal Photoreception in Early Vertebrates
2010
[49]
논문
The Evolution of Trichromatic Color Vision by Opsin Gene Duplication in New World and Old World Primates
http://genome.cshlp.[...]
1999-07-01
[50]
논문
Trichromacy in Australian Marsupials
2002-04
[51]
웹사이트
Mammalia – Palaeos
http://www.palaeos.c[...]
[52]
논문
Modern mammal origins: evolutionary grades in the Early Cretaceous of North America
1989-07
[53]
논문
A Jurassic mammal from South America
2002-03
[54]
논문
The oldest platypus and its bearing on divergence timing of the platypus and echidna clades
2008-01
[55]
논문
Molecules, morphology, and ecology indicate a recent, amphibious ancestry for echidnas
2009-10
[56]
웹사이트
Appendicular Skeleton
http://courses.washi[...]
[57]
논문
Review of the early allotherian mammals
http://www.app.pan.p[...]
[58]
웹사이트
Mammaliaformes—Palaeos
http://palaeos.com/V[...]
2005-05-21
[59]
웹사이트
http://www.palaeos.c[...]
[60]
웹사이트
Mammalia: Spalacotheroidea & Cladotheria – Palaeos
http://palaeos.com/v[...]
[61]
웹사이트
Metatheria – Palaeos
http://www.palaeos.c[...]
2007-04-19
[62]
논문
The Mongolian Late Cretaceous Asiatherium , and the early phylogeny and paleobiogeography of Metatheria
1996-09-19
[63]
웹사이트
Oldest Marsupial Fossil Found in China
http://news.national[...]
National Geographic News
2003-12-15
[64]
웹사이트
Didelphimorphia – Palaeos
http://palaeos.com/v[...]
2013-10-15
[65]
논문
Angolan ichnosite in a diamond mine shows the presence of a large terrestrial mammaliamorph, a crocodylomorph, and sauropod dinosaurs in the Early Cretaceous of Africa
2017-04
[66]
웹사이트
Family Peramelidae (bandicoots and echymiperas)
http://animaldiversi[...]
[67]
웹사이트
Species is as species does... Part II
http://lancelet.blog[...]
2005-12-12
[68]
웹사이트
Marsupials
http://paleo.amnh.or[...]
[69]
논문
Epipubic bones in eutherian mammals from the Late Cretaceous of Mongolia
1997-10
[70]
논문
An analysis of epipubic bone function in mammals using scaling theory
1989-08
[71]
논문
A Jurassic eutherian mammal and divergence of marsupials and placentals
2011-08
[72]
웹사이트
Eomaia scansoria: discovery of oldest known placental mammal
http://www.evolution[...]
[73]
논문
The placental mammal ancestor and the post-K-Pg radiation of placentals
2013-02
[74]
논문
Hypaxial motor patterns and the function of epipubic bones in primitive mammals
2003-01
[75]
논문
Epipubic bones in eutherian mammals from the late Cretaceous of Mongolia
1997-10
[76]
뉴스
Why we don't lay eggs
https://www.newscien[...]
[77]
논문
Viruses and the placenta: the essential virus first view
https://escholarship[...]
2016-01
[78]
웹사이트
Eutheria – Palaeos
http://www.palaeos.c[...]
2007-04-19
[79]
논문
The earliest known eutherian mammal
2002-04
[80]
논문
A large therian mammal from the Late Cretaceous of South America
2024-02-03
[81]
논문
Skeleton of a Cretaceous mammal from Madagascar reflects long-term insularity
https://research.mon[...]
2020-04-29
[82]
서적
Paleontology and Geology of the Upper Jurassic Morrison Formation: Bulletin 36
https://nmdigital.un[...]
New Mexico Museum of Natural History and Science
2006
[83]
논문
A multivariate approach to infer locomotor modes in Mesozoic mammals
2015-02-24
[84]
논문
A Late Jurassic digging mammal and early mammalian diversification
2005-04
[85]
논문
A Mesozoic gliding mammal from northeastern China
http://doc.rero.ch/r[...]
2006-12
[86]
논문
New materials of Argentoconodon fariasorum (Mammaliaformes, Triconodontidae) from the Jurassic of Argentina and its bearing on triconodont phylogeny
[87]
논문
A new family of primitive mammal from the Mesozoic of western Liaoning, China
2001-05
[88]
논문
Large Mesozoic mammals fed on young dinosaurs
http://doc.rero.ch/r[...]
2005-01-13
[89]
서적
The Age of Dinosaurs in Russia and Mongolia
Cambridge University Press
2003
[90]
논문
Carnivorous dental adaptations in tribosphenic mammals and phylogenetic reconstruction
2007-03-29
[91]
논문
Two possibly aquatic triconodont mammals from the Early Cretaceous of Morocco – Acta Palaeontologica Polonica
https://www.app.pan.[...]
1995
[92]
논문
An extraordinary fossil captures the struggle for existence during the Mesozoic
[93]
논문
Mammaliaform extinctions as a driver of the morphological radiation of Cenozoic mammals
2021-07
[94]
논문
A large therian mammal from the Late Cretaceous of South America
2024-02-03
[95]
논문
Resolution of the early placental mammal radiation using Bayesian phylogenetics
2001-12
[96]
논문
Retroposed elements as archives for the evolutionary history of placental mammals
http://biology.plosj[...]
2006-04
[97]
논문
Retroposon analysis and recent geological data suggest near-simultaneous divergence of the three superorders of mammals
2009-03
[98]
논문
Mosaic retroposon insertion patterns in placental mammals
2009-05
[99]
논문
Impacts of the Cretaceous Terrestrial Revolution and KPg extinction on mammal diversification
2011-10
[100]
웹사이트
Scientists map elephant evolution
http://news.bbc.co.u[...]
2008-08-11
[101]
웹사이트
Historical perspective (the Dynamic Earth, USGS)
http://pubs.usgs.gov[...]
2007-06-24
[102]
웹사이트
Cretaceous map
http://www.scotese.c[...]
2007-04-18
[103]
웹사이트
Overview
http://www.palaeos.c[...]
[104]
논문
Secondary structure and patterns of evolution among mammalian mitochondrial 12S rRNA molecules
1996-10
[105]
논문
Compensatory substitutions and the evolution of the mitochondrial 12S rRNA gene in mammals
1995-11
[106]
서적
Molecular Evolution
Sinauer Associates
[107]
논문
The delayed rise of present-day mammals
2007-03-29
[108]
논문
The Interrelationships of Placental Mammals and the Limits of Phylogenetic Inference
2016-02
[109]
논문
Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary
2007-06
[110]
논문
Early origins of modern birds and mammals: molecules vs. morphology
1999
[111]
논문
Fossil Evidence for a Late Cretaceous Origin of 'Hoofed' Mammals
1996-05-24
[112]
논문
Primate Origins: Implications of a Cretaceous Ancestry
2007
[113]
서적
PRIMATE ORIGINS: Adaptations and Evolution
[114]
논문
The Fossil Record of North American Mammals: Evidence for a Paleocene Evolutionary Radiation
1999-03-01
[115]
논문
Quantitative Analysis of the Timing of the Origin and Diversification of Extant Placental Orders
2001-06-01
[116]
논문
Independent origins of middle ear bones in monotremes and therians
2005-02
[117]
논문
The Mammary Gland and Its Origin During Synapsid Evolution
2002-07
[118]
논문
The Origin of Lactation as a Water Source for Parchment-Shelled Eggs
2002-07
[119]
웹사이트
Lactating on Eggs
http://nationalzoo.s[...]
[120]
논문
Washington's pesticide panel: process and product
1993-06
[121]
서적
Mammal-like reptiles and the origin of mammals
Academic Press
[122]
논문
A New Tritylodontid from the Upper Jurassic of Xinjiang, China
[123]
논문
The dentary of Wareolestes rex (Megazostrodontidae): a new specimen from Scotland and implications for morganucodontan tooth replacement
https://ora.ox.ac.uk[...]
[124]
웹사이트
Evolutionary Trends In The Mammalian Digestive System
https://campus.murra[...]
2019-09-30
[125]
논문
Invited review: ruminant ecology and evolution: perspectives useful to ruminant livestock research and production
2010-04
[126]
논문
The Evolution of Stomach Acidity and Its Relevance to the Human Microbiome
2015-07-29
[127]
서적
Digestive Physiology and Metabolism in Ruminants
Springer Netherlands
1980
[128]
논문
Transmission modes of the mammalian gut microbiota
2018-10
[129]
논문
Experimental Evidence for Adaptation to Species-Specific Gut Microbiota in House Mice
2019-07
[130]
논문
Role of the Gut Microbiome in Vertebrate Evolution
2018-04-24
[131]
웹사이트
Study suggests gut bacteria helped shape mammalian evolution
https://phys.org/new[...]
2019-09-30
[132]
논문
Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia
2016-10
[133]
논문
A study on the skeleton of ''Diademodon''
[134]
서적
Mammal-like reptiles and the origin of mammals
Academic Press
[135]
논문
Cranial anatomy of the cynodont reptile ''Thrinaxodon liorhinus''
https://www.biodiver[...]
[136]
웹사이트
"''Thrinaxodon:'' The Emerging Mammal"
http://news.national[...]
National Geographic Daily News
2012-08-26
[137]
논문
Une piste avec traces de soies épaisses dans le Trias inférieur a moyen de Lodève (Hérault, France): ''Cynodontipus polythrix nov. gen., nov. sp.'' les Cynodontes en France
[138]
간행물
"''Cynodontipus'': A procolophonid burrow – not a hairy cynodont track (Middle-Late Triassic: Europe, Morocco, Eastern North America)"
http://gsa.confex.co[...]
Geological Society of America
2012-03
[139]
논문
Palaeoneurological clues to the evolution of defining mammalian soft tissue traits
2016-05
[140]
뉴스
Getting to the Root of Fur
https://www.national[...]
2014-10-10
[141]
서적
Animal physiology: Adaptation and environment
https://archive.org/[...]
Cambridge University Press
[142]
서적
Comparative Animal Physiology
Saunders College
[143]
논문
Limb posture in early mammals: Sprawling or parasagittal
https://www.app.pan.[...]
2006
[144]
서적
Perspectives of Biophysical Ecology
[145]
논문
Do extraordinarily high growth rates in Permo-Triassic dicynodonts (Therapsida, Anomodontia) explain their success before and after the end-Permian extinction?
2010-07-26
[146]
웹사이트
Ancestry of mammalian 'warm-bloodedness' revealed
https://www.scienced[...]
Society of Vertebrate Paleontology
2015-10-29
[147]
서적
Predatory Dinosaurs of the World
https://archive.org/[...]
Simon and Schuster
[148]
논문
Monotreme Cell-Cycles and the Evolution of Homeothermy
1988
[149]
논문
The evolution of nasal turbinates and mammalian endothermy
1992
[150]
논문
The Evolution of Endothermy in Mammals and Birds: From Physiology to Fossils
1995-10
[151]
논문
The Evolution of Endothermy in the Phylogeny of Mammals
1978-01
[152]
논문
A caseian point for the evolution of a diaphragm homologue among the earliest synapsids: Early synapsid origin of a diaphragm homologue
2016-12
[153]
논문
The postcranial skeleton of African cynodonts
[154]
서적
Vertebrate Life
https://archive.org/[...]
Prentice-Hall
[155]
뉴스
Mammals' large brains evolved for smell
https://www.bbc.co.u[...]
2011-05-22
[156]
뉴스
Mammals' bodies outpaced their brains right after the dinosaurs died
https://www.sciencen[...]
2022-03-31
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com